



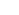
Dingo Úc có phải là chó nhà (Canis Familiaris) - Một trường hợp gây tranh cãi về phân loại (Phần II)
17/03/2023* Tên bài báo đã được người dịch đặt lại.
Tác giả: Bradley P. Smith và cộng sự
Phân biệt dingo với các loài chó khác
Jackson và cs. (2017) tập trung vào “... mức độ tương thích di truyền đang tồn tại” rõ ràng giữa dingo và chó nhà, đồng thời lập luận rằng vì chó dingo (1) chia sẻ dòng gen liên tục với chó nhà và có thể dễ dàng lai với chó nhà, (2) được cho là đã được con người du nhập vào lục địa Úc và (3) trùng lặp về mặt hình thái với chó nhà, do đó dingo nên được phân loại là một giống chó nhà. Điều này phản ánh sự nhầm lẫn và tranh cãi có tính chất toàn cầu về việc đặt tên cho các loài có nguồn gốc hoang dã và được nuôi (xem phần sau). Thật vậy, những lập luận này không phản ánh đầy đủ tính phức tạp của vấn đề phân biệt các loài của chi canis và không xem xét đầy đủ về tính độc đáo của loài dingo. Dưới đây chúng tôi phác thảo những lý do bổ sung tại sao chó dingo có thể và nên được phân biệt với chó nhà.
So sánh kiểu hình
Ngoại hình chung. Con dingo trưởng thành trung bình nặng khoảng 15 kg, cao 55 cm tính từ vai và dài 123 cm từ mũi đến chóp đuôi (Corbett 2001a; Smith 2015a). Smith (2015b) đã phác thảo một số đặc điểm được coi là điển hình (hoặc dấu hiệu nhận biết) của chó dingo thuần chủng có thể được sử dụng để phân biệt chó dingo với hầu hết các giống chó vì chúng không phải là đặc điểm chung của chó nhà (xem Sanchez-Villagra và cs. 2016). Ví dụ, cơ thể của dingo dài hơn chiều cao của nó; lồng ngực dài và mở rộng ra phía sau; đầu là phần rộng nhất của cơ thể (tức là nó có ngực hẹp và không mở rộng xuống dưới khuỷu tay); hộp sọ cao ở sau tai và tai hướng về phía trước của chẩm; đôi mắt xếch, hình quả hạnh nhân và nằm xiên; và đuôi thường cụp thấp, không cong ra phía sau và có một tuyến mùi hương ở trên đuôi. Để biết ví dụ về kiểu hình dingo 'điển hình', hãy tham khảo Hình 1.

Hình 1. Ví dụ về ngoại hình dingo điển hình. Bức ảnh mô tả một con chó đực từ đảo K’gari-Fraser (Queensland) của John Williams.
Màu lông (ví dụ: màu lông đồng nhất, các mảng màu trắng) có thể cung cấp một số dấu hiệu về sự khác biệt giữa dingo thuần chủng và dingo lai chó nhà (Newsome & Corbett 1985; Jones 1990; Corbett 2001a,b). Màu sắc biểu thị sự lai tạp (đặc trưng di truyền của chó nhà) là màu vện, loang lổ, đốm và kiểu yên ngựa (ví dụ: chó chăn cừu Đức), được kiểm soát bởi các đột biến hiện đại dường như là gắn liền với quá trình thuần hóa và chọn lọc nhân tạo, và sẽ không xuất hiện ở dingo 'thuần chủng', không có tổ tiên là chó nhà hiện đại (Cairns và cs. 2011; Smith 2015a). Có một nhận định chung, cả trong tài liệu khoa học và công chúng nói chung, rằng những con dingo có màu vàng (hoặc màu gừng) đồng nhất với các mảng màu trắng ở bàn chân. Mặc dù điều này là phổ biến, thì người ta cũng tìm thấy những tấm da cũ có màu sắc khác ở những vùng hẻo lánh trong Thế kỷ 19, bao gồm cả các cá thể có màu đen với tứ chi màu nâu, cá thể có lông trên lưng sẫm màu (màu lông chồn) và màu trắng (Crowther và cs. 2014). Sự biến đổi của bộ lông khiến việc phân biệt dingo với những con lai với chó nhà một cách chắc chắn trở nên khó khăn. Nhìn chung, việc so sánh kiểu hình với chó nhà là khó khăn (Elledge và cs. 2006, 2008; Newsome và cs. 2013), đặc biệt còn do sự biến đổi lớn về ngoại hình của chó nhà (Crowther và cs. 2014; Drake & Klingenberg 2010). Do đó, thường có sự trùng lặp giữa các giống chó nhà và các loài của chi canis hoang dã (Crowther và cs. 2014; Smith và cs. 2018).
Hình thái hộp sọ. Sự tách biệt về hình thái của dingo khỏi các loài chó khác đã được ghi nhận từ lâu (Zeuner 1963; Epstein 1971; Macintosh 1975; Barker & Macintosh 1979). Dingo thường có hộp sọ nhỏ hơn chó sói (ví dụ: chiều dài hộp sọ của loài dingo nhỏ hơn khoảng 15% so với loài sói nhỏ Ấn Độ C. lupus pallipes) và có sự khác biệt về hộp sọ (Gollan 1982; Corbett 2001a; Crowther và cs. 2014; Parr và cs. 2016; Geiger và cs. 2017) và hình dạng tai trong (Schweizer và cs. 2017). So với loài sói, loài dingo cũng có hình dạng hộp sọ đồng dạng, đặc trưng bởi khuôn mặt ngắn hơn và rộng hơn (Geiger và cs. 2017). Ngoài ra, chó dingo cho thấy sự khác biệt rõ ràng về hình thái hộp sọ với chó bản địa ở Thái Lan và Indonesia (Gollan 1982; Corbett 1985; Gonzalez 2012). Dingo cũng khác về mặt hình thái so với chó biết hát New Guinea (C. hallstromi) với chiều rộng gò má tương đối nhỏ hơn (Koler-Matznick et al. 2003; Crowther và cs. 2014). So với chó nhà, dingo 'thuần chủng' có mõm dài hơn (Newsome và cs. 1980; Newsome & Corbett, 1982), răng nanh và răng hàm lớn hơn, răng nanh dài hơn và mảnh hơn một chút, hốc thính giác lớn hơn (Newsome và cs. 1980; Newsome & Corbett 1982; Corbett 2001a), kích thước não/cơ thể lớn hơn (Smith và cs. 2018) và hộp sọ phẳng hơn với gáy (Newsome và cs. 1980; Newsome & Corbett 1982; Geiger và cs. 2017) và xương đỉnh đầu lớn hơn (Gollan 1984; Jones 1990). Hình dạng bên trong tai của dingo giống chó sói hơn là giống nhiều giống chó khác (Schweizer và cs. 2017). Tuy nhiên, sự khác biệt về mặt hình thái của chó dingo với chó nhà vẫn là một vấn đề vì những lý do sau:
1. Có nhiều biến thể hình thái về hình dạng hộp sọ ở chó nhà hơn so với tất cả các loài trong toàn bộ Bộ ăn thịt (Drake & Klingenberg 2010);
2. Chó lai dingo-chó nhà và dingo có thể khó phân biệt về mặt hình thái (Newsome & Corbett 1982; Jones 1990; Elledge và cs. 2008; Newsome và cs. 2013; Parr và cs. 2016);
3. Sự khác biệt hình thái và phân tử của dingo và chó nhà dựa trên các mẫu động vật sau khi tiếp xúc với người châu Âu, hoặc những động vật bị nuôi nhốt (Newsome và cs. 1980; Newsome & Corbett, 1982; Elledge và cs. 2006, 2008; Wilton và cs. 1999; Wilton , 2001); do đó, các mẫu có thể có dòng dõi thể hiện sự lai cận giới;
4. Một số giống chó, đặc biệt là chó chăn gia súc của Úc, cho thấy bằng chứng về sự lai giống với dingo trong quá khứ (Arnstein và cs. 1964), có khả năng gây khó khăn trong việc cung cấp các đặc điểm xác định rõ ràng và nhất quán để phân biệt dingo với chó nhà trong quá khứ (Newsome và cs. 1980);
5. Dingo thể hiện sự biến đổi hình thái theo không gian và thời gian, chủ yếu là về kích thước (Corbett 2001a,b; Radford và cs. 2012; Claridge và cs. 2014; Colman 2015);
6. Có bằng chứng di truyền của ít nhất hai quần thể phân loài dingo, có thể là kết quả của nhiều lần du nhập vào Úc (Cairns & Wilton 2016; Cairns và cs. 2017). Các quần thể phân loài này cũng cho thấy một số bằng chứng về sự tách biệt về hình thái (Colman 2015), do đó làm mờ thêm các đặc điểm nhận dạng có thể có giữa dingo và chó nhà.
Crowther và cs. (2014) đã nhận ra những khó khăn này trong việc cung cấp các đặc điểm nhận dạng hình thái đáng tin cậy để phân biệt dingo với chó nhà. Do đó, họ đã định loài cho dingo không phải bằng cách sử dụng sự tách biệt hoàn toàn của các ký tự trong giản đồ phân tích thành phần chính và phân tích lưỡng biến, mà bằng cách định loài cho dingo như một cụm chặt ở trong sự biến đổi của chó nhà thuần hóa. Parr và cs. (2016) cũng chỉ ra một nhóm dingo trong phạm vi rộng hơn của chó hoang và chó nhà trong toàn bộ hình thái hộp sọ. Mặc dù chồng lên nhau, nhưng những con dingo có chiều rộng vòm miệng tương đối lớn hơn, mõm dài hơn, chiều cao hộp sọ ngắn hơn và các đường gờ thái dương của hộp sọ rộng hơn (Crowther và cs. 2014) so với chó nhà. Hơn nữa, hộp sọ của dingo không hiển thị bất kỳ sửa đổi hoặc thay đổi cực đoan nào do quá trình thuần hóa (và tỷ lệ cận huyết cao kéo theo điều này) chẳng hạn như mắt tròn hơn và hướng về phía trước, mõm ngắn lại và răng mọc quá nhiều sau đó, giảm kích thước của xương đỉnh đầu và xương chóp gáy, và bọng nước màng nhĩ giảm (Jones 1990) (Hình 2).
Hình 2. Tái tạo hộp sọ 3-D của một con dingo (phía dưới) và một con chó nhà thả vườn (trên cùng), làm nổi bật sự khác biệt về hình thái hộp sọ được đề cập trong văn bản. Hộp sọ chó đã được thu nhỏ để phù hợp với chiều dài của hộp sọ dingo để tạo điều kiện so sánh đặc điểm hình dạng. Con dingo (đực) được thu thập từ Trạm Minburra ở Nam Úc. Con chó (cái, 14,5 kg) được thu thập vào năm 1981 từ Cao nguyên Victoria (Bộ sưu tập của Evan Jones).
Crowther và cs. (2014) cũng đã giải quyết vấn đề về sự không chắc chắn liên quan đến tổ tiên của loài dingo bằng cách sử dụng các mẫu vật trước khi có sự tiếp xúc với người châu Âu, cũng như các mẫu vật từ các vùng của Úc trước năm 1900, nơi người châu Âu định cư thưa thớt. Mặc dù lai tạo có thể đóng một vai trò trong việc tạo ra các kiểu màu lông (Newsome & Corbett 1985; Cairns và cs. 2011; Crowther và cs. 2014), nhưng dường như nó đã góp phần rất ít vào việc thay đổi hình dạng hộp sọ của loài chó dingo. Parr và cs. (2016) đã sử dụng phương pháp tiếp cận hình thái học-hình học 3-D để định lượng hình thái sọ (xương) tổng thể của chó dingo, chó nhà và chó lai dingo-chó nhà. Họ đã sử dụng chó dingo và chó lai dingo-chó nhà được nhân giống để nghiên cứu thử nghiệm (Catling và cs.1992; Newsome & Corbett 1982), cũng như tuyển chọn chó nhà và chó hoang (bắt từ hoang dã). Chó lai Dingoes-chó nhà và dingo hình thành các cụm riêng trong không gian hình thái của chi canis và không thể phân biệt được về mặt thống kê, trong khi chó nhà được phân bố rộng rãi trên khắp không gian hình thái. Điều này có thể một phần là do chó nhà được lai tạo có chọn lọc để giữ lại những đặc điểm kiểu hình đôi khi cực đoan, thường là tính trạng lặn về mặt di truyền, dẫn đến sự cố định tính trạng lặn đồng hợp tử nhiều hơn trong bộ gen. Những con dingo hoang dã trải qua quá trình chọn lọc tự nhiên trên một nguồn gen rộng hơn, duy trì các kiểu hình tính trạng trội và kiểu gen dị hợp tử (Parr và cs. 2016).
Hầu hết các loài chó trên thế giới sống trong mối quan hệ bán tự quản bởi con người (Barnett & Rudd 1983; Gompper 2014). Trong tình trạng phổ biến nhất này, kiểu hình của chó nhà có một số đặc điểm chung với dingo. Tuy nhiên, kiểu hình này biến đổi nhiều hơn so với kiểu hình của dingo - tức là chúng vẫn có thể biểu hiện tai cụp, bộ lông đốm/loang lổ - tùy thuộc vào tổ tiên của giống chó nhà. Chó bản địa ở Đông Nam Á có thể có kiểu hình bề ngoài tương tự như dingo, nhưng có thể phân biệt với dingo trên một loạt các biến số hình thái, kiểu hình và di truyền (Corbett 1985; Gonzalez 2012; Cairns & Wilton 2016).
Jackson và cộng sự đã ghi nhận quan sát của Parr rằng “... hình thái của dingo thuần chủng và lai tạp trùng nhau rất nhiều nên không thể phân biệt con lai một cách đáng tin cậy với dingo dựa trên số đo sọ não” (Jackson và cs 2017; Parr và cs. 2016, tr. 210-211). Mặc dù điều này đúng một phần, nó đòi hỏi sự cẩn trọng do những gì Parr trên thực tế mới chỉ ra rằng dingo và chó lai dingo-chó nhà không thể phân biệt dựa trên phân tích hình thái của hình dạng tổng thể của hộp sọ. Điều này bao gồm việc kiểm tra các điểm mốc được sử dụng bởi Newsome và Corbett (Newsome và cs. 1980, Newsome & Corbett 1982 và Corbett (2001a, b) không thể phân biệt hình dạng sọ của dingo và con lai. Tuy nhiên, những phát hiện này không loại trừ việc dingo có những hình thái độc đáo trong các vùng cụ thể của hộp sọ.
Hơn nữa, Parr và cộng sự không chỉ ra rằng phép lai làm thay đổi hình dạng của động vật con cháu, mà là sự kiện lai tạo qua các thế hệ liên tiếp (tức là F1 = dingo × chó giống, F2 = dingo × F1, F3 = dingo × F2, v.v.), làm cho các con lai biểu hiện hình thái dingo. Điều này được giải thích thông qua dòng dingo về cơ bản đại diện cho dòng gen "kiểu hoang dã", ít nhất là so với những con chó giống, thường được lai ngược để "cố định" các đặc điểm kiểu hình mong muốn (ít nhất là đối với con người). Nói cách khác, 'chó giống' đại diện cho các kiểu gen hẹp hơn, thường là lặn, mà kiểu hình của chúng không được biểu hiện sau thế hệ F2 của các con lai với dingo 'kiểu hoang dã'. Điều này hỗ trợ khẳng định của Parr và cộng sự về sự pha trộn giữa dingo và chó nhà không loại trừ hình thái của dingo bản địa (Parr và cs. 2016).
Nguồn gốc và bản sắc di truyền
Các nghiên cứu về di truyền học cho thấy chó dingo Úc phù hợp với hệ phát sinh loài của chi Canis và có quan hệ họ hàng gần nhất với chó nhà ở vùng phía nam của Đông Á (Savolainen và cs. 2004; vonHoldt và cs. 2010; Oskarsson và cs. 2011; Ardalan và cs. 2012; Sacks và cs. 2013; Wang và cs. 2016; Cairns&Wilton 2016). Điều này rất phù hợp với các phân tích hình thái học dựa trên các phép đo được thực hiện trên sọ chó dingo, chó sói và chó nhà (Gollan 1982). Mặc dù có nguồn gốc từ tổ tiên giống chó nhà (like dog), nhưng theo một loạt các dấu hiệu di truyền, chó dingo khác biệt về mặt di truyền với các giống chó hiện đại, bao gồm cả chó bản địa châu Á và có chung một số dấu hiệu di truyền với chó sói châu Á (Savolainen và cs. 2004; vonHoldt và cs. 2010; Sacks và cs. 2013; Freedman và cs. 2014; Cairns & Wilton 2016; Wang 2016, Zhang và cs.2018). Dữ liệu về ty thể và nhiễm sắc thể Y chỉ ra rằng chó dingo có quan hệ mật thiết với chó nhà Đông Nam Á (Savolainen và cs. 2004; Oskarsson và cs. 2011; Ardalan và cs. 2012; Sacks và cs. 2013).
Có một loạt các dấu hiệu di truyền có thể phân biệt dingo với chó nhà, bao gồm các dấu hiệu ty thể và nhiễm sắc thể Y (Savolainen và cs. 2004; vonHoldt và cs. 2010, Oskarsson và cs. 2011; Ardalan và cs. 2012; Sacks và cs. 2013; Cairns & Wilton, 2016; Cairns và cs. 2017), cũng như các DNA vi vệ tinh (microsatellite) và đa hình nucleotide đơn hạt nhân (SNP). Kể từ những năm 1990, xét nghiệm DNA dựa trên kỹ thuật “microsatellite” đã được sử dụng để phân biệt dingo, dingo có tổ tiên chó nhà và chó hoang. Ban đầu dựa trên 14 microsatellite, 23 marker (bao gồm hai marker đánh dấu chèn - xóa [indel]) hiện nay vẫn đang được sử dụng (Wilton và cs. 1999; Wilton 2001; Elledge và cs. 2008; Newsome và cs. 2013; Stephens và cs. 2015). Những marker này được chọn vì sự khác biệt về phân bố alen giữa quần thể dingo tham chiếu và quần thể của chó nhà lai tham chiếu (Wilton và cs. 1999; Wilton 2001).
Một chỉ trích chính đối với các phương pháp dựa trên microatellite này để phân biệt dingo với dingo có một phần tổ tiên chó nhà là thiếu mẫu tham chiếu thời kỳ trước khi người châu Âu đến (Crowther và cs. 2014). Thay vào đó, mẫu tham chiếu dingo bao gồm một tập hợp dingo do các khu bảo tồn, sở thú hoặc bộ sưu tập mẫu vật có mô tả kiểu hình dingo không xác định do ‘chuyên gia’ cung cấp. Đây là các trường hợp điển hình vì tại thời điểm nghiên cứu không có mô tả chính thức về các đặc điểm cơ thể để làm cơ sở xác định dingo (Corbett 2001b; Wilton 2001; Elledge và cs. 2008) cho đến khi Crowther và cộng sự cung cấp một cơ sở dựa trên các đặc điểm cơ thể của các mẫu vật dingo thế kỷ 18 và 19 (Crowther và cs. 2014).
Tuy nhiên, nghiên cứu di truyền ban đầu đã phát hiện ra rằng quần thể chó nhà và dingo tham chiếu là khác biệt về mặt di truyền, và gợi ý rằng sự pha tạp gen của chó nhà trong quần thể chó dingo tham chiếu là thấp (Wilton và cs. 1999; Wilton 2001, Zhang và cs. 2018). Những tiến bộ trong công nghệ đã cải thiện phương pháp ban đầu bằng cách áp dụng phân nhóm Bayes, giảm nhẹ sự phụ thuộc vào mẫu tham chiếu chó dingo hoặc chó nhà, được xác định trước, cỡ mẫu nhỏ (Stephens 2011; Stephens và cs. 2015). Gời đây, việc lập mô hình để suy ra tổ tiên của dingo bằng cách sử dụng một quần thể tham chiếu được xác định trước, nhưng cũng cho phép so sánh giữa hàng trăm (hoặc hàng nghìn) con dingo và dingo có kiểu gen đã biết với một vài tổ tiên chó nhà. Tuy nhiên, 23 marker được sử dụng hiện tại cung cấp phạm vi giới hạn của bộ gen, vì vậy các xét nghiệm trong tương lai lý tưởng là nên kết hợp các mẫu chó dingo thời kỳ đầu hoặc tiền Châu Âu và lấy mẫu di truyền mật độ cao hơn để theo dõi sự lai tạp.
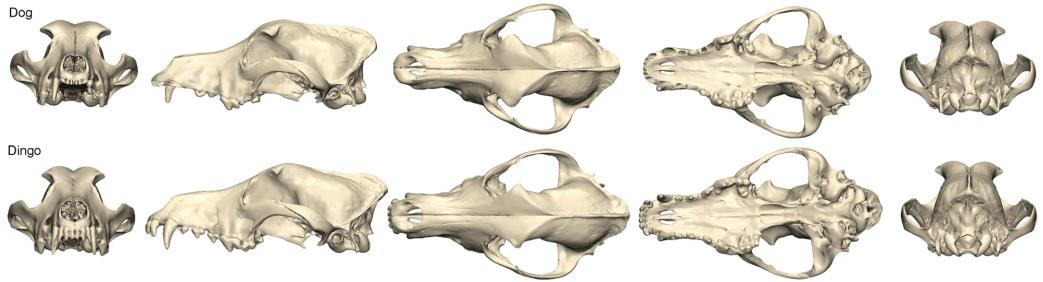
Ngày nay, dựa trên những tiến bộ đặc biệt gần đây trong công nghệ ADN, việc giải trình tự toàn bộ bộ gen nhân và phân tích bằng cách sử dụng bảng đa hình đơn nucleotide (SNP) mật độ cao trên toàn bộ bộ gen cung cấp các công cụ mạnh mẽ để phân biệt dingo với các loài khác thuộc chi canis (vonHoldt và cs. 2010; Freedman và cs. 2014; Cairns 2015). Những phân tích này cho thấy dingo có dòng dõi chó nhà cổ xưa, phân nhánh sớm (vonHoldt và cs. 2010; Freedman và cs. 2014; Cairns & Wilton 2016; Wang và cs. 2016, Zhang và cs. 2018). Ví dụ, phân tích thành phần chính dựa trên SNP toàn bộ bộ gen giúp phân biệt dingo với các quần thể của chi chó khác: chó bản địa và các giống chó hiện đại, cũng như chó sói (Hình 3). Những dữ liệu này chỉ ra rằng dingo (và chó New Guinea) tạo thành một quần thể riêng biệt khác với chó sói và chó nhà hiện đại (Hình 4).
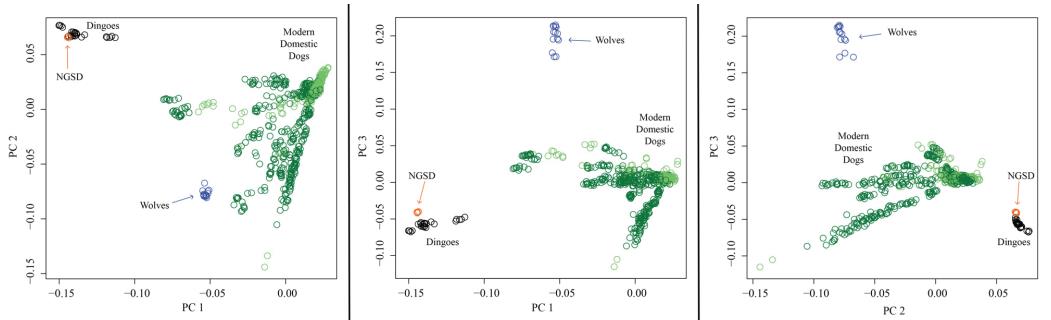
Hình 3. Sự biến đổi di truyền giữa chó dingo, chó biết hát New Guinea, chó sói và chó nhà hiện đại dựa trên dữ liệu SNP trên toàn bộ bộ gen được thể hiện bằng cách sử dụng phân tích thành phần chính (PCA) đại diện cho 841 con chó (xanh lá cây), 24 con chó dingo (đen), 5 New Guinea singing (màu cam), 14 con sói (màu xanh lam). Trong số những con chó nhà có 292 con chó giống (màu xanh lá cây nhạt) và 549 con chó bản địa (màu xanh lá cây đậm). Chúng tôi định kiểu gen cho từng mẫu chó này bằng chip Illumina CanineHD 170 SNP (dữ liệu từ Cairns 2015 và Shannon và cs. 2015). Chúng tôi đã phân tích dữ liệu kiểu gen trong PLINK 1.9 (Chang et al. 2015) bằng cách hợp nhất các bộ dữ liệu và loại bỏ các SNP bị thiếu trong hơn 10% mẫu. Tổng số marker SNP còn lại là 166.019. Chúng tôi đã thực hiện PCA trong PLINK 1.9 và hiển thị kết quả bằng cách sử dụng R (v 3.2.1). PC1 chiếm 49,2% biến dị, PC2 chiếm 20,7% biến dị và PC3 chiếm 10,8% biến dị 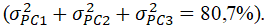
Các nghiên cứu từ sớm sử dụng các phương pháp di truyền để điều tra nguồn gốc của dingo đã khẳng định rằng dingo ở Úc đã hình thành một quần thể đồng nhất, có thể là kết quả của một đợt nhập cư duy nhất (Savolainen và cs. 2014). Tuy nhiên, Cairns và Wilton đã sử dụng toàn bộ bộ gen của ty thể và quan sát thấy rằng có hai dòng dõi ty thể khác biệt về mặt phát sinh gen ('đông nam' và 'tây bắc') ở dingo (Cairns và Wilton 2016). Hai dòng này có sự phân bố được chia nhỏ về mặt địa lý, trong đó một dòng hầu như chỉ giới hạn ở đông nam Úc (đông nam) và dòng còn lại ở các khu vực còn lại của Úc (miền trung; phía bắc và phía tây; phía tây bắc). Mặc dù ít rõ ràng hơn, dữ liệu nhiễm sắc thể Y-cũng cho thấy bằng chứng về cấu trúc địa lý (Ardalan và cs. 2012; Sacks và cs. 2013; Cairns và cs. 2017) và gợi ý rằng loài dingo có thể bắt nguồn từ ít nhất hai lần di cư, tạo thành hai quần thể cơ bản mà sau đó trải qua sự pha trộn thiên vị giới tính, với chó đực di chuyển nhiều hơn chó cái. Nguồn gốc địa lý cuối cùng của dòng chó đực và chó cái vẫn chưa được giải quyết hoàn toàn, nhưng thực tế là mỗi dòng trong số các dòng ty thể và nhiễm sắc thể Y đều có chung với chó biết hát New Guinea cho thấy rằng một số cuộc di cư có thể đã xảy ra thông qua cầu đất liền nối giữa New Guinea và Australia (bị ngắt kết nối hoàn toàn vào khoảng 7.000 năm trước; Torgersen và cs. 1988) - có hoặc không có sự can thiệp của con người (Cairns & Wilton 2016; Cairns và cs. 2017).
Hiện tại, có hai đến ba quần thể di truyền của loài dingo: đông nam, tây bắc và đảo K’gariFraser (Cairns&Wilton 2016; Cairns và cs. 2017). Những con dingo ngày nay thể hiện một số biến thể về hình thái và kích thước trên khắp lục địa gần như phù hợp với sự biến đổi môi trường và sự phân chia về mặt di truyền (Colman 2015), dẫn đến cái gọi là các dạng dingo 'núi cao', 'sa mạc' và 'nhiệt đới'. Tuy nhiên, khi so sánh các marker di truyền, ADN nhiễm sắc thể Y và ty thể, DNA vệ tinh và dữ liệu SNP trên toàn bộ bộ gen, dingo nhìn chung được nhận diện là một quần thể khác biệt, phân nhánh sớm với tất cả các loại chó nhà cũng như các loài của chi canid hoang dã khác.
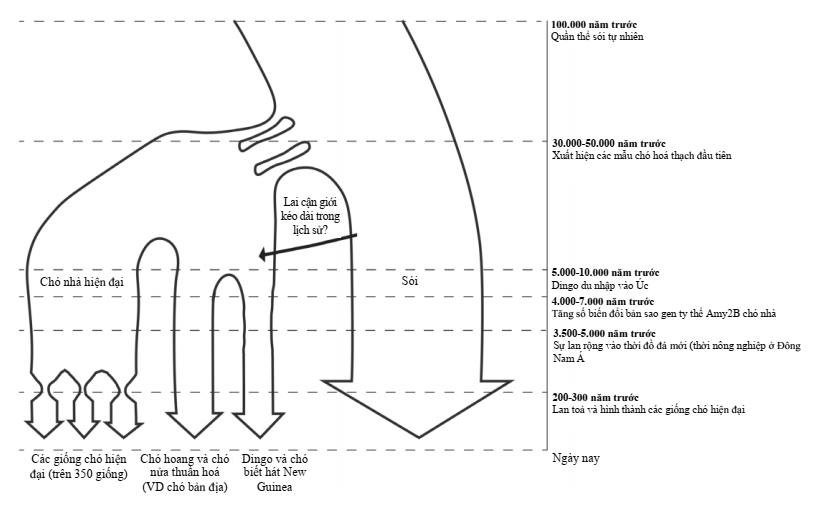
Hình 4. Tóm tắt mối quan hệ tiến hóa giữa chó sói, chó dingo và chó nhà hiện đại. Hình này là tổng hợp của các mô hình nguồn gốc chó khác nhau (vonHoldt và cs. (2010), Cairns & Wilton (2016), vonHoldt & Driscoll (2016)). Dingo và các dòng chó cổ xưa khác như chó biết hát New Guinea tạo thành một dòng riêng biệt tách biệt với những con chó nhà hiện đại đã trải qua nhiều thế hệ chọn lọc nhân tạo liên tục.
So sánh hành vi
Sinh sản. Mùa sinh sản phân biệt rõ ràng chó dingo với các nhóm chó nhà tiến hóa gần đây dựa vào nguồn thức ăn do con người tạo ra (Lord và cs. 2013). Giống như tất cả các loài của chi canis hoang dã, dingo đực và cái có kiểu sinh sản theo mùa (một lứa mỗi năm), có liên quan đến, nhưng không hoàn toàn được quyết định bởi quang kỳ (Lord và cs. 2013; Smith 2015a, Cursino và cs. 2017). Đối với loài chó dingo, mùa sinh sản thường diễn ra vào mùa thu (tháng 2 đến tháng 6), với quá trình đẻ con diễn ra vào mùa đông (tháng 6 đến tháng 8) (Catling 1979; Catling và cs. 1992; Thomson 1992a; Jones & Stevens 1988; Catling và cs. 1992; Smith 2015a). Dingo có khả năng sinh sản trong năm tuổi đầu tiên của chúng vào mùa sinh sản, và đặc biệt là khi các nhóm đàn bị phá vỡ (Wallach và cs. 2009), nhưng hầu hết không sinh con cho đến khi chúng được hai tuổi (Breckwoldt 1988; Catling và cs. 1992; Jones & Stevens 1988). Động dục có thể thay đổi, xảy ra từ một đến bốn tuổi (Jones & Stevens 1988). Ngược lại, chó nhà và chó bản địa có thể sinh sản hai lần một năm (vào bất kỳ thời điểm/mùa nào trong năm), thường bắt đầu động dục sau mỗi 6-9 tháng (Boitani và cs. 2006, Majumder & Bhadra 2015). Trong một số trường hợp, chó bản địa chỉ sinh sản mỗi năm một lần, mặc dù thời gian giao phối dài (lên đến 5 tháng) so với các loài chó hoang dã (Pal 2001, 2011). Các nghiên cứu về chó bản địa chỉ ra rằng chó đực và chó cái sinh sản trong năm đầu tiên (Boitani và cs. 1995); tuy nhiên, điều này có thể khác nhau tùy thuộc vào kích thước của con chó (Lord và cs. 2013).
Hành vi lãnh thổ và tổ chức bầy đàn. Nói chung, chó dingo thể hiện các hành vi lãnh thổ và bầy đàn giống như đã được mô tả đối với các loài của chi chó hoang dã khác, một số hành vi này không phổ biến trong các nhóm chó nhà. Trong hầu hết các trường hợp, một nhóm gia đình hoặc đàn dingo có thể duy trì một lãnh thổ hoặc khu vực riêng biệt bao quanh nơi ở và thường trung thành với địa điểm này từ năm này sang năm khác (Thomson 1992a,b; Corbett 2001a). Các nhóm thường chứa một cặp đực và cái sinh sản chính hoặc ưu thế (bố mẹ) và con cái của chúng ở các độ tuổi khác nhau, với bầy từ 3 đến 12 cá thể; những cá thể không liên quan hiếm khi được phép tham gia đàn (Thomson 1992a; Corbett 2001a). Để so sánh, chó hoang/chó bản địa có xu hướng sống theo nhóm lớn, nhưng trong hầu hết các trường hợp không hình thành các nhóm xã hội riêng biệt hoặc liên kết cặp riêng biệt, hoặc lãnh thổ xác định và chúng phụ thuộc vào thức ăn do con người tạo ra hơn là con mồi hoang dã (Berman & Dunbar 1983; Pal và cs. 1999; Boitani và cs. 2006). Tuy nhiên, các cộng đồng chó bản địa và chó hoang rất khác nhau, với sự chênh lệch về mức độ hành vi xã hội được quan sát trên các địa điểm nghiên cứu khác nhau (Cafazzo và cs. 2010).
Giao tiếp bằng âm thanh. Sủa có ở tất cả các loài của chi chó, mặc dù có sự khác biệt giữa các phân loài. Dingo sủa trong bối cảnh bị đe dọa (ví dụ: tương tác kích động ở cự ly gần và khi có người lạ tiếp cận), và như một đoạn của điệp khúc hú (Déaux&Clarke 2013). Tuy nhiên, dingo không sủa trong các tương tác giao tiếp, ngay cả khi mức độ kích thích cao, chẳng hạn như trong các tương tác vui vẻ (Byosiere và cs. 2018). Điều này trái ngược với chó nhà, chúng dường như sủa trong mọi tình huống, trong các tương tác kích động, khi bị báo động, lúc cho ăn hoặc khi chúng bị cô lập về mặt xã hội (Feddersen-Petersen 2000; Yin & McCowan 2004; Lord và cs. 2009). Điều quan trọng là, tiếng sủa thường được xuất hiện khi chúng chơi đùa và là một tín hiệu quan trọng để làm trung gian cho các trò chơi tương tác (Bekoff 1972; Yin & McCowan 2004; Horowitz 2009; Smuts 2014). Một điểm khác biệt nữa giữa dingo và hầu hết các giống chó khác là dingo phát ra tiếng hú như một cách để bảo vệ lãnh thổ của chúng (Corbett 2001a). Trong khi một số giống chó có thể và phát ra tiếng hú (ví dụ: husky Siberia), hầu hết thì không (Bradshaw&Nott 1995), và chức năng của nó vẫn chưa được biết. Điều quan trọng là chó bản địa ở Ý và Ấn Độ sử dụng tiếng sủa để bảo vệ lãnh thổ chống lại đồng loại, nhưng không tru dài (Boitani & Ciucci 1995; Pal 2015). Do đó, trong khi hầu hết các hành vi giao tiếp được bảo tồn cao trong chi Canis, thì có những khác biệt quan trọng giữa dingo và chó nhà.
Nhận thức. Mặc dù có mối quan hệ gần gũi giữa chó sói, chó nhà và dingo, nhưng chúng khác nhau trong cách tiếp cận để giải quyết vấn đề. Giống như chó sói (Frank 1980; Frank & Frank 1985, 1987), dingo giỏi giải quyết các vấn đề không mang tính xã hội hơn chó nhà. Ví dụ, có nghiên cứu (Smith và Litchfield 2010) đã chỉ ra rằng dingo giải quyết vấn đề về không gian thành công hơn nhiều so với chó nhà (Pongrácz và cs. 2001, 2005, 2008), thể hiện độ trễ ngắn hơn và ít lỗi hơn, cũng như không có dấu hiệu thất bại hoặc nhầm lẫn. Hơn nữa, giống như loài sói (Fox, 1971; Frank 1980; Mech 1991), dingo đã thể hiện các hành vi giải quyết vấn đề phức tạp như sử dụng công cụ trong điều kiện nuôi nhốt (Smith và cs. 2012) và đã được ghi nhận là phản ứng trước cái chết của một con cùng loài trong một quần thể hoang dã theo cách mà trước đây chỉ được quan sát thấy ở các loài có trí khôn khác như linh trưởng, một số động vật biển có vú và voi (Appleby và cs. 2013). Có kết luận rằng một trong những điểm khác biệt chính giữa hành vi của chó nhà và chó sói là việc chó nhà sẵn sàng giao tiếp bằng mắt với con người và sử dụng cách nhìn như một phương thức giao tiếp (Miklosi và cs. 2003). Trong khi đó, ngay cả sau khi xã hội hóa mạnh mẽ, những con sói dường như không quan tâm đến các tín hiệu giao tiếp của con người hoặc tham gia vào các tương tác giao tiếp với con người (Miklósi 2007). Johnston và cs. (2017) đã điều tra sở thích giao tiếp bằng mắt với con người của dingo. Trái ngược với loài sói (Nagasawa và cs. 2015), dingo bắt đầu giao tiếp bằng mắt với con người, nhưng trong thời gian ngắn hơn so với chó nhà và chúng có xu hướng tránh giao tiếp bằng mắt trong thời gian dài (Johnston và cs. 2017). Chúng tôi đồng ý với đánh giá, dingo không có động cơ như chó nhà để bắt đầu hoặc tìm cách giao tiếp bằng mắt với con người hoặc để duy trì nó (Smith và Litchfield 2013). Johnston và cs. (2017) cho rằng những con dingo có thể đã phát triển một số động lực để bắt đầu các tương tác đầu tiên bằng mắt với người, nhưng không phát triển mong muốn duy trì tiếp xúc này vì điều này xảy ra ở giai đoạn muộn của quá trình thuần hóa chó nhà.
Nguồn: https://doi.org/10.11646/zootaxa.4564.1.6
Người dịch: Phạm Thu (P.TTKHQS)
Bài viết liên quan






Khoa học & công nghệ



